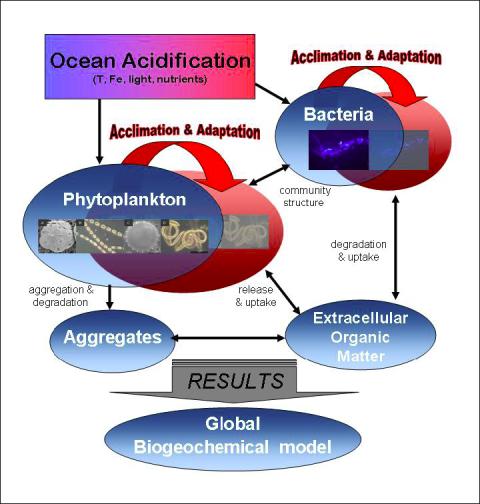
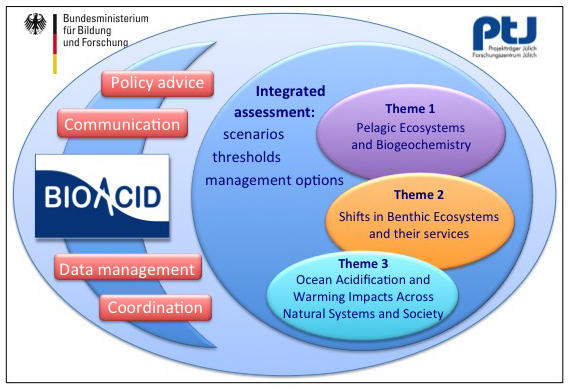
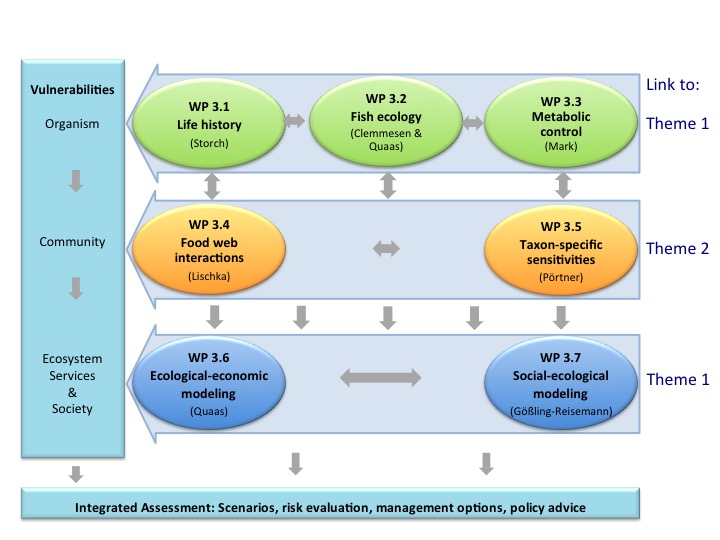
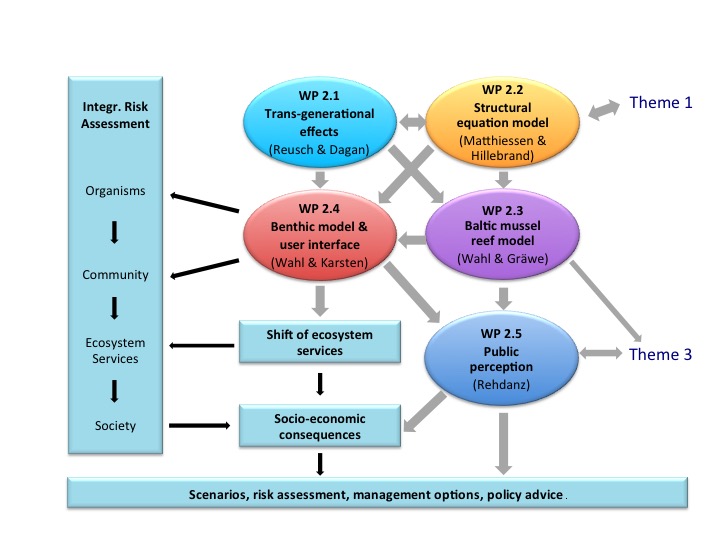
Lead Proponent: Dr. Dirk de Beer (Max-Planck-Institute for Marine Microbiology, Bremen
We would like to predict the state of the future oceans. Which organisms will be present? How will the seas function in global element cycling, and how can oceans be used as a resource for humans? The importance of the seas for humanity as a source of food, for recreation and even as aesthetic and spiritual resource can not be overestimated.
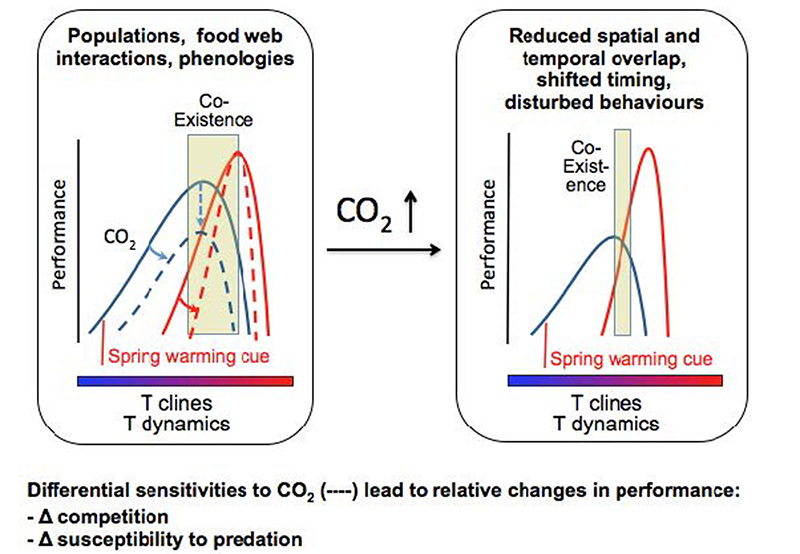
Ocean acidification (OA) by rising atmospheric carbon dioxide (CO2) concentrations can be predicted reasonably well. Conversely, the effects on marine diversity and biological functions are very hard to predict. Since often organisms have long life spans, estimating how organisms and communities will adapt to the future pH decrease is difficult and cannot be addressed by short-term laboratory experiments. For example, in a short-term experiment, the initial stress responses can blur possible acclimatization mechanisms, and furthermore, evaluating the evolutionary implications would also be challenged by short incubation.
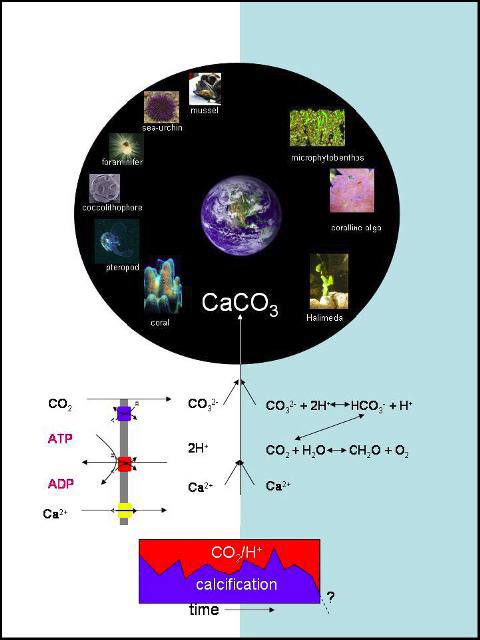
Moreover, the complexity of OA is increased by other environmental changes than pH. For instance, OA predicts surface temperature, increased evaporation and therefore salinity changes, a change in metal and nutrient chemistry and concentrations, and combined or separate, each change will effect species fitness and community productivity. Biogenic reefs, which depend on calcification, may be particularly vulnerable. Decreased oversaturation may endanger reef building and increase erosion. Reefs deserve special attention as they are hotspots of biodiversity and productivity, and are important spawning and nursing grounds.
We will study ecologically and economically important reef communities from tropical, temperate and sub-arctic CO2 enriched habitats to assess crucial questions on the long-term effects of ocean acidification. At such sites, species should be adapted and acclimatized to high seawater pCO2 or have disappeared, and therefore, the community reflects potential directions of future changes to new equilibriums in the future oceans. We will focus on benthic organisms that have key roles in ecosystem engineering: the reef builders and primary producers.
The chosen sites serve as our windows and models for the future oceans. Whereas the views are obscure and narrow, what we will see is a realistic approximation. We will use the sites as natural laboratory, and by studying acclimatization and adaptation of microbes, micro- and macro algae, invertebrates and corals along gradients of carbonate chemistry and nutrients. We will transplant micro and macro-organisms (corals, algae) in order to study their physiology and acclimatization potential. Laboratory measurements will be included, on site and in the institutes, to separate out effects of specific environmental variables and to study experimental evolution and adaptation capacity of model organisms over longer time scales. Finally, we will talk to stakeholders, the local communities and others directly depending on the reefs. We hope to learn how these ecosystem users will cope, or plan to cope, with the expected changes, and how they will estimate the magnitude of economic losses or gains under different scenarios of acidification.
General objectives:
The studies we propose encompass three levels: 1) organism physiology, 2) biodiversity and 3) ecosystem functioning. We will determine the impact of OA on metabolic functions, and determine if OA increases energy demands. Knowledge on which organisms are affected, and which not, will be used to predict changes in diversity. Finally, the measured decrease or increase in diversity and changes in metabolic performance will lead to predictions in the changes of key ecosystem functions. A decreased pH, altered carbonate chemistry and nutrient levels may not necessarily affect the key ecological process rates of primary production and degradation. Diversity may change in response to OA, but the major biogeochemical processes may be sustained with a modified species assemblage (Kelly, Barott et al. 2011). For example, calcifying organisms may be replaced by non-calcifiers. We will investigate the underlying physiological mechanism for pH dependent competiveness of species, with a focus on pH/pCO2 dependence of their energy budgets. We will study if and how adaptation and acclimatization to enhanced pCO2 occurs, by studying natural habitats that are since centuries enriched in CO2. An important benefit for studying the physiology of organisms from such natural CO2 sites is that this biota has been exposed to rising CO2 for longer duration, and therefore we avoid studying stress responses, and rather assess acclimatization and impact.
We plan to address the following basic hypotheses:
Hypothesis 1: Adaptation and acclimatization leads to communities that are more resistant against low pH, their action spectrum has a lower pH optimum. The specific carbonate chemistry of selected sites has existed for centuries to millennia. This is crucial for assessing long-term effects of ocean acidification on community structures and ecosystem functioning.
Hypothesis 2: Adaptation and acclimation of calcifying organisms to elevated seawater pCO2 requires surplus energy. Energy demands impact growth and thus reduce competitive abilities. Ensuing increased vulnerability of reefs will lead to significant reductions in economic revenue generated from these ecosystems.
Hypothesis 3: Low pH is accompanied by increased nutrients and trace metals that can protect against low pH. We will assess physiological responses with an emphasis on growth rate, gene expression, and primary and new production estimates. Furthermore, responses will be assessed with single cell and bulk approaches, as often a response could be variable even within a group (genus) of organisms.
These hypotheses will form the basis of the research on the arctic, temperate and tropical reefs.
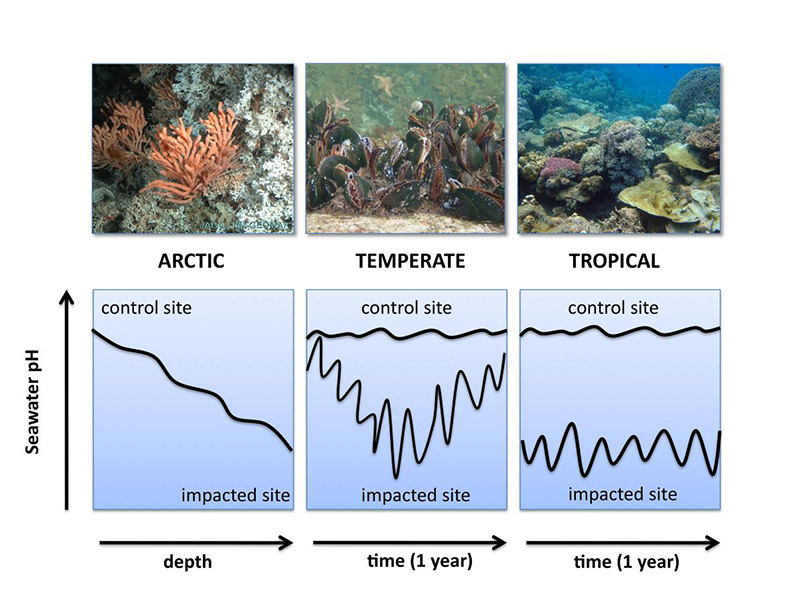
Consortium 4 studies reef communities in arctic, temperate and tropical waters.
Graph: BIOACID
Research approaches:
We will study a variety of benthic organisms: corals, macrophytes, microphytobenthos, bivalves, bacteria and archaea. Comparisons of warm water and cold water corals in mesocosm experiments will be included. The integration of the consortium will be strengthened by joint campaigns to the field sites.
Study sites will be the CO2 seeps in Papua New Guinea (PNG, tropical), bivalve reefs in the Western Baltic and North Sea (temperate), and cold water corals reefs along the Norwegian shelf (sub-arctic). The biological diversity of the PNG site is among the highest in the world (Fabricius, Langdon et al. 2011). There are several well defined gas seeps (sulfidic and pure CO2), that maintain gradients in habitat from acidic to seawater, harbour a variety of sediment types (pH 6 to 8.2, iron-rich clay, siliceous and calcareous sands). The first field surveys on the reef off PNG indicate pronounced shifts in the abundance of different functional groups of benthic primary producers (corals, macrophytes, coralline algae) along the pH-gradient. The western Baltic is characterized by pronounced seasonal oscillations in pCO2, with average surface gradients in pCO2 between 400 and 700 μatm, and summer averages of >1000 μatm at some locations (Thomsen, Gutowska et al. 2010). CaCO3 under saturation is a common phenomenon. We will compare differently impacted sites in the Baltic (low pCO2, high pCO2) with sites in the North Sea (low pCO2) and study, whether populations have adapted to high pCO2. Mesocosms in Kiel will be used to study physiology of cold water corals retrieved during cruises from sub-arctic sites (in situ pH ~7.8).
The field work will consist of sampling for diversity, rate measurements and experimental transplantation of living specimen to assess their capability to acclimatise to environmental changes. In addition, artificial settlement plates will be exposed within pCO2 gradients to study colonization, diversity and productivity of developing communities. Laboratory studies will accompany field work to determine action spectra for pH and nutrients and to study energy budget allocation, as well as adaptation capacity of model species.
Cruises to PNG are planned for February 2013 and August 2014. Fieldwork in the Baltic and North Sea will be planned in a flexible manner.
Biogeochemistry and microbial processes:
We will determine physico-chemical habitat parameters (e.g. pCO2, pH, nutrients, metals, O2, salinity, temperature). Different spatial resolutions are required to determine variability within the site using microsensors on a micro-scale (10-100 mm), and by geochemical analyses of sediments and porewater. Loggers for pH, Temperature (T) and dissolved oxygen (O2) will be moored at sites to assess temporal variations. In addition, carbon and nitrogen cycling, such as carbon (C) fixation, degradation of organic matter (aerobic and anaerobic respiration), and N2-fixation will be measured along gradients by whole water bottle incubations. The main functions may not be influenced by OA, but the microbial communities performing these tasks may differ. Furthermore, single cell approaches using a high-resolution nanometre scale secondary ion mass spectrometry (nanoSIMS) will help assess the cell to cell variation in metabolic activity (i.e. C and N2 fixation) within a given community.
Physiology:
Effects of pCO2, temperature and nutrients on organism performance will be assessed. Special attention will be paid to energy budgets, as these might be modulated and decrease a group’s competiveness, particularly in calcifying organisms. Energy budgets will be determined on an organismal and cellular level to determine costs for pH homeostasis, calcification, stress and immune responses. Carbonate structures (stability, composition, microstructure) will be analyzed, and related to erosion of reefs. Effects of pCO2 on key processes in primary production of calcifying and non-calcifying algae will be determined. Comparative studies on tropical and cold water corals will determine their pH dependant fitness and growth rates. Action spectra with regard to pH and nutrients of key microbial processes will be determined. Laboratory experiments will be conducted in Bremen, Kiel and Sylt laboratories that have facilities for CO2 perturbation experiments, and during field campaigns on site.
Diversity and adaptation:
We will determine community compositions by molecular methods along gradients of pH and nutrients, and determine the main parameters driving community shifts using mathematical methods. Physiology and community structure will be linked to water and sediment chemistry. Evolutionary biology experiments on model invertebrates (bivalves) will investigate whether high CO2 tolerance observed in populations from high pCO2 sites is an inheritable trait, allowing us to determine the extent of past selection events on genotype x environment interactions (Hoffman and Sgro, 2011). The studies will focus on low dispersive taxa, such as brooding corals, seagrass and ascidians.
Socio-economic consequences of ocean acidification on reefs:
Economic impacts of ocean acidification on coral reefs will be performed through a choice experiment (Brander, Rehdanz et al. 2009). An assessment of potential changes in mollusc fishery and aquaculture revenues will be performed using partial-equilibrium analysis, global up scaling will be attempted using the climate-economy models IMPACT and FUND (Narita, Rehdanz et al. 2011). This study is now placed in WP 5.4 and will there be described in detail.
Integration and management:
The consortium is conceptually unified by the addressing the same hypotheses, and practically by implementing similar research approaches in the three different habitats:
The physico-chemical habitat descriptions (pCO2, pH, temperature, salinity, nutrients, metals, POC, currents) will be a joint effort and the same techniques will be used.
We will determine the microbial community structures (incl. diversity, richness) and functions (primary production, mineralization, N2-fixation), as well as the shifts therein along pH gradients.
At all three sites settlement experiments will be done, using colonization tiles, to study the microbial and macrobiota (animal and plant) diversity, succession and competition. Furthermore the tile approach can assess impact on biomass and CaCO3 production. Physiological responses of the ecosystem engineering species (bivalves and corals) will be determined following long-term acclimatization. The energy budgets will help explain competitive strength of the key species, responsible for the structural integrity of reefs. Studies of the carbonate skeletons of bivalves, corals and coralline algae will be performed in the three climatic sites.
We will perform joint studies at each field site, share methods and data, exchange students, and present and discuss each others results in regular meetings.

Laboratory experiments accompany the fieldwork.
Photo: Armin Form, GEOMAR
Output for the BIOACID consortium:
Data on the effects of pH change on primary production of benthic communities and general element cycling will become available for foodweb studies/modeling. The effects of enhanced metabolism of calcifying organisms, and ensuing reduced growth rates of e.g. bivalves are essential for bivalve fisheries and aquacultures.
We expect that we will be able to predict if and what form reefs will be able to survive the pH decrease. Especially important here is whether calcifying key-players will be outcompeted by non-calcifying populations. E.g. will corals as primary producers be replaced by macrophytes, or will mussel beds be overgrown by algae.
Finally, the cruises to New Guinea will host social and economic scientists from Consortium 5. Using the habitat descriptions we will construct future ecosystem scenarios, which will be used to investigate how the coastal populations will adapt to the consequences of pH decrease.